There is a high number of theoretically possible homozygous and stable chromosome combinations. Under the presumption that the substitutions are restricted to homoeologous rye and D genome chromosomes the number can be calculated from the general formula 2n. In this case n is 7, that is the number of chromosome pairs in each of the participating genomes. The distribution of different possible chromosome combinations will be the following :
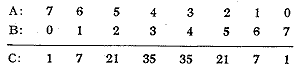
A represents the number of rye chromosome pairs and B the number of D genome chromosome pairs (or vice versa). The sum of A and B is seven, that is a full genome. C represents the number of different possible chromosome combinations. E.g. there are seven different combinations of six rye and one D genome chromosome, since each of the seven rye chromosomes theoretically can be substituted by its homoeologous D genome chromosome. In addition to these 128 different R-D combinations there is the possibility of substitutions between the wheat genomes. A line with e.g. four pairs of rye chromosomes does not necessarily have three pairs of D genome chromosomes. It could have more, since D genome chromosomes can substitute for chromosomes of the A and B genomes. This means that the production of secondary hexaploid triticale opens the possibility of a far reaching reorganisation of the chromosomal and genomic composition. Hexaploid triticale crossed to bread wheat seems to be the most suitable combination for the selection of different chromosome types. Maybe the optimal chromosome composition of hexaploid triticale has not yet been isolated.
In practice, however, there are indications that a specific F1 between hexaploid triticale and bread wheat is limited in its chromosome substitution possibilities (MERKER 1975). There are two plausible explanations for this : One is gene interaction and the other is lack of homoeology between rye and wheat chromosomes. A pair of D genome chromosomes in a certain line may lack a specific gene that is essential for the successful replacement of a rye homoeologue, or have a specific gene that makes it impossible as a substitute for a rye homoeologue. Cultivated rye, Secale cereale, is differentiated from species of wild rye by reciprocal translocations involving either two or three chromosome pairs (see KRANZ 1963). F1:s between different wheat cultivars reveale the presence of a varying number of reciprocal translocations (see LARSEN 1973). These structural differences disturb the homoeology between rye and wheat chromosomes and counteract full compensation and balanced genotypes when chromosomes are substituted.